|
|||||||||||||||||||||||||||||||||||||||||||||||||||

![]()
|
Phyllotaxie – Développements Simulations numériques Principales
hypothèses expliquant l'arrangement doré
des feuilles ou fleurs et approches mathématiques. De 1800 à 1950, les chercheurs ont proposés des
modèles tentant d'expliquer les lois observées de la phyllotaxie. Plus les
études avancent et plus le modèle physico-biochimique se complexifie au point
d'avoir besoin de recourir à la simulation numérique sur ordinateurs, et cela pour
découvrir leur comportement et connaitre les résultats. C'est Turing qui est le premier à se lancer. Mais
les moyens informatiques de l'époque sont dérisoires au regard de ceux de maintenant. Cette page est faite pour tous ceux qui
voudraient toucher du doigt le domaine de la phyllotaxie explicative.
Autrement-dit, il s'agit d'une approche simplifiée (simpliste). Toute erreur
d'interprétation serait de mon fait. |
|
|
||
|
Mathématisation |
D'Arcy Thompson (1860-1948) est le premier
biomathématicien. En 1917, il publie: On
Growth and Form (réédité
en 1942) Il
prétend que ses prédécesseurs (comme Julius Sachs)
ont sous-estimé le rôle de la physique
et de la mécanique dans la constitution des formes des organismes vivants. En
phyllotaxie, il retrouve la suite
de Fibonacci. Il montre
qu'on peut passer d'une forme d'une espèce à la forme d'une espèce proche par
certaines transformations
géométriques (anamorphose). Il disait: Il existe encore une autre manière – c'est à
Henri Poincaré que nous la devons –
de considérer la fonction des mathématiques, et de bien comprendre pourquoi
ses lois et ses méthodes sous-tendent nécessairement la science physique tout
entière. Tout phénomène naturel, si simple qu'il paraisse, est en réalité
composite, et chaque action, chaque effet visible est en réalité la somme
d'un nombre indéfini d'actions sous-jacentes. C'est ici que s'exprime le
pouvoir tout particulier des mathématiques: celui de combiner et de
généraliser. |
|
|
Théorème fondamental de la
phyllotaxie |
Ce théorème (Adler – 1974) vise à clarifier la confusion due à
l'observation d'hélices multiples.
(Illustration en couleur par Wolfram). Ci-contre,
la représentation d'une cellule dans un |
|
|
Modélisation numérique |
La
représentation géométrique de la théorie de la diffusion d'un inhibiteur (Schoute) est difficile sinon impossible. Seule, une
simulation dynamique peut témoigner de l'action d'inhibition au fur et à
mesure de la naissance des primordia. Ces modèles aboutissent à des résultats
qui confirment ce que crée la nature, témoignant que les paramètres pris en
compte ont toutes les chances d'être les bons. Veen-Lindenmayer (1978)
reprend le modèle géométrique de Van Iterson(1907)
et introduit le concept de gradient
chimique. Un réel progrès, car ce modèle prend en compte le facteur temps. Thorney en 1975; Young en 1978;
etc. |
|

|
|
||
|
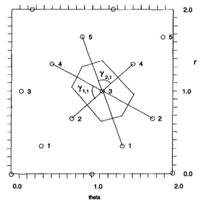
Principes
de base de la simulation Les
graines apparaissent à la surface d’un cercle central appelé apex. Chaque
nouvelle graine apparait à la surface de ce cercle à l’endroit le plus
dégagé, celui où la distance avec les graines déjà présentes est la plus
grande. Une fois
apparues, les graines s’éloignent toutes du centre à la même vitesse qui ne
dépend que de la distance au centre. Ici, sur
l'illustration, le motif en spirale
est bien reproduit et on trouve le couple (13,21) ce qui correspond bien à la
propriété attendue. |
|
|

|
Simulation numérique |
|
|
|
Les
scientifiques tentent de donner une
explication à cette corrélation magique
entre botanique et mathématique. La
question reste ouverte en 2015. |
Voir État d'avancement en 2015. |
|
|
Un modèle
géométrique qui tenterait d'expliquer les mouvements au sein du méristème (lieu où s'exerce la croissance par division
cellulaire) serait trop succinct. Alan Turing, célèbre mathématicien s'attaque à
ce problème. Il a émis l'hypothèse que chaque primordium émet autour de lui
une substance inhibitrice empêchant la formation d'autres primordia à sa
périphérie. Hélas son décès laissera ce travail inachevé. En 1996, Stéphane Douady et Yves Coudier reprennent la simulation de l'apparition des
primordia à partir du critère d'Hofmeister. Ils ont
mis en évidence que l'angle de divergence dépendait fortement d'un facteur
appelé G. |
En 1868,
Hofmeister remarque que: Les primordia apparaissent un à un, dans des
intervalles réguliers; Dans la zone de naissance, les primordia se
déplacent dans le plus grand espace disponible en évitant ainsi au maximum
les primordia existants; et Les organes existants se déplacent radialement,
avec le tissu de croissance. Ces observations ont conduit les scientifiques à
imaginer des modélisations reproduisant ce mode de croissance. Les
explications possibles
|
|
|
Seule une
modélisation du phénomène dynamique pourrait donner les clés, les paramètres
gouvernant la croissance. Cependant, en tenant compte seulement
sans tenir compte
le modèles atteignent déjà une formidable complexité
mathématique. Compte
tenu des choix nécessaires, aucune modélisation jusqu'à présent n'a donné
entière satisfaction. Elles n'expliquent pas tous les cas et surtout les cas
d'irrégularités. Pourtant, chacune conduisent à l'organisation
"en or" observée sur les plantes. |
De nombreuses tentatives de modélisations
existent. Au niveau des méristèmes, il y a plusieurs
couches (L1, L2 et L3), chacune avec des comportements différents. Aucune explication biologique précise ne permet de
conclure sur la manière dont se forme la configuration spiralée. Au moins quatre modèles ont été émis, qui se
complètent plus ou moins:
|
|
|
Un exemple: le modèle de Gierer et Meinhardt (GM – ci-contre) du type activateur-inhibiteur
est le plus apprécié des modèles de formation des formes biologiques. Il fait
appel aux dérivées partielles (A et H sont les concentrations de l'activateur
et de l'inhibiteur). La mise
en place des équations et leur
résolution sont du niveau post-bac. Les
solutions ne sont généralement pas analytiques (pas de fonctions explicites).
Le recours à l'analyse par ordinateur
s'impose. |
Idée de la
complexité des modélisations Un autre
type de formulation
|
|
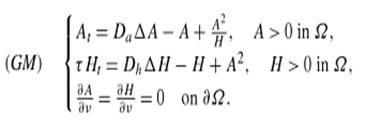
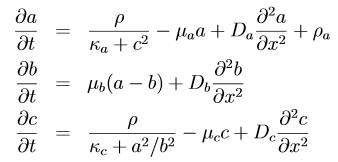
|
|
||
|
Actuellement,
ce qui est le plus important, c'est de comprendre comment les cellules du
méristème (MAC) communiquent pour
déclencher celles qui vont créer le nouveau
primordium. Les trois
théories indiquées ci-contre sont, en fait, complémentaires. Elles supposent
que le point de création d'une nouvelle feuille est déterminé par la
diffusion chimique d'un champ de croissance
qui peut être inducteur ou inhibiteur. Il
n'existe aucune modélisation physiologique confirmée par l'expérience qui
soit universellement acceptée par les chercheurs. |
Trois écoles:
|
|
Selon Handbook
of Plant Science – Volume 2 – Page 102 – Ediror
Keith Roberts – 2007 – 1300 pages
|
Les simulations de modèles physico-chimiques sont
concordantes avec une majorité des caractéristiques de la structure des
plantes, notamment pour les parastiques en
nombres de Fibonacci. Avec le développement de la puissance de calcul des ordinateurs,
les modèles peuvent être de plus en plus sophistiqués et, désormais, ils
tiennent compte d'un grand nombre de paramètres physico-biochimiques. Néanmoins, les chercheurs sont toujours à la recherche de la
théorie unique qui concilierait toutes les hypothèses crédibles connues
actuellement. |
![]()
|
Retour |
|
|
Voir |
|
|
Sites |
|
|
Cette page |
http://villemin.gerard.free.fr/aScience/Botanique/PhylNume.htm |
![]()